Italiano |
Discussion |
|||||||
![]()
With the research carried out in the mountain and hilly part of the catchment basin of Rio Santa Lucia it has been possible to assess 669 taxa, of which 629 were species, 35 subspecies, 2 varieties and 3 hybrids, belonging to 364 genera and 101 families (Tab. 4). The Dicotyledones with 70 families, 266 genera and 486 entities were dominant over the other systematic groups.
FAMILIES
GENERA
SPECIES
SUBSPECIES
VARIETIES
HYBRIDS
Pteridophyta 13
16
23
2
-
-
Gymnospermae 3
3
4
-
-
-
Dicotyledones 70
266
456
28
2
-
Monocotyledones 15
79
146
5
-
3
Total 101
364
629
35
2
3
Tab. 4. Taxa flora of Rio Santa Lucia.
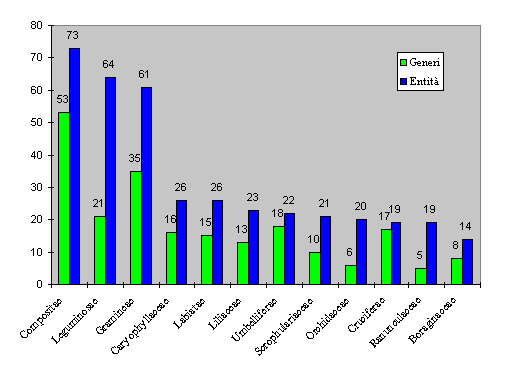
The families with the greatest number of entities (Fig. 2 and Tab. 5) are in the order the Compositae (73), the Leguminosae (64) and the Gramineae (61). An interesting point about the Umbelliferae and the Cruciferae is that their number of genera (respectively 18 and 17) is very near their number of entities (22 and 19).
Fig. 2. Families with the greatest number of genera and entities.
Families Genera
Species
Subspecies
Hybrids
Total
Compositae 53
70
3
-
73
Leguminosae 21
60
4
-
64
Gramineae 35
60
1
-
61
Caryophyllaceae 16
25
1
-
26
Labiatae 15
23
3
-
26
Liliaceae 13
23
-
-
23
Umbelliferae 18
19
3
-
22
Scrophulariaceae 10
21
-
-
21
Orchidaceae 6
14
3
3
20
Cruciferae 17
18
1
-
19
Ranunculaceae 5
17
2
-
19
Boraginaceae 8
14
-
-
14
Tab. 5. Families with the greatest number of taxa.
Tab. 6 shows a comparison of the floristic richness of the studied area with that of the nearby areas of Pixinamanna (Arrigoni 1964), Pantaleo-Gutturu Mannu-Punta Maxia (Camarda & al. 1993), Monte Tamara (Ballero & al. op. cit.) and Monte Arcosu (Mossa & al. 1996). In considering the flora of Pixinamanna we did not include any taxa lower than subspecies. This reduced the entities from 552 to 467. From a comparison of the different florae, it was found that the Rio Santa Lucia catchment basin is floristically poorer that all other areas, except for Pantaleo-Gutturu Mannu-Punta Maxia. These data are justified by the remarkable difference in area, by the nature of the substrates and by the relative homogeneity of the examined territory compared to the areas of Monte Tamara, Monte Arcosu and Pixinamanna. The only really comparable area by size and territorial homogeneity was the Pantaleo-Gutturu Mannu-Punta Maxia area, that is remarkably poorer in terms of numbers and their relation with the area.
FLORA Area (Kmq)
Entity
Entity/Kmq
Genera
Gen./Kmq
Families
Fam./Kmq
Rio Santa Lucia 73
669
9.2
364
5.0
101
1.4
Monte Tamara (BALLERO & al. op. cit.) 25
472
18.9
294
11.8
78
3.1
Monte Arcosu (MOSSA & al. 1996) 32
520
16.3
303
9.5
90
2.8
Pixinamanna (ARRIGONI 1964) 40
467
11.7
269
6.7
n.s.
-
Pantaleo (CAMARDA & al. 1993) 120
593
4.9
338
2.8
90
0.8
Tab. 6. Floristic richness.
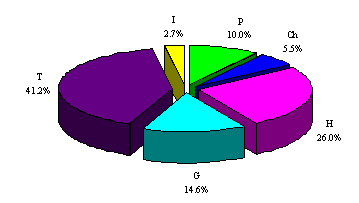
The data relating to the biological spectrum (Fig. 3) basically confirm the Mediterranean nature of the area (T=41.2%) and the high degree of wooded cover (P=10%). The high percentage of geophytes (G=14.6%) seems related to the anthropic use of the land, especially to the practice of fires and sylvo-pastoral types of activities. The percentage of hydrophytes, that are mainly located along the torrents and in the neighbourhood of the springs, is significant (I=2.7%).
Fig. 3. Biological spectrum of the flora of the Rio Santa Lucia catchment basin.
From a comparison of the biological spectrum (Tab. 7) with that of Sardinia (Bocchieri 1995) no particular disagreement emerges. The only differences are in the therophytes values compared to the florae of Pixinamanna (Arrigoni 1964) and Pantaleo-Gutturu Mannu-Punta Maxia (Camarda & al. 1993). The therophyte percentage values observed for these florae were significantly higher (10.8 and 7 percentage points), though there are no important landscape justifications except that of an exposure to the sea for the Pixinamanna area and of an inclusion of agricultural areas in the Pantaleo-Gutturu Mannu-Panta Maxia area. As regards recent surveys on Monte Tamara and Monte Arcosu, from a comparison with the flora of Rio Santa Lucia, the observed values seem to agree except for the hydrophytes. This is attributable to the geomorphological and geological nature of the two systems. In fact both present very few watercourses and springs. Moreover Monte Tamara is made up of calcareous rocks that present deep karst phenomena.
P
Ch
H
G
T
I
Rio Santa Lucia 10.0
5.5
26.0
14.6
41.2
2.7
Pixinamanna (Arrigoni 1964) 9.0
5.0
19.0
15.0
52.0
0.0
Pantaleo (Camarda & al. 1993) 9.3
3.7
26.7
11.4
48.2
0.7
M. Tamara (Ballero & al. op. cit.) 10.0
5.0
26.0
14.0
45.0
0.0
M. Arcosu (Mossa & al. 1996) 10.4
5.0
25.6
14.0
43.6
1.4
Sardegna (Bocchieri, op. cit.) 8.8
8.1
28.1
12.1
39.9
3.0
Tab. 7. Comparision of biological spectra.
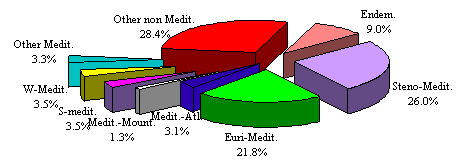
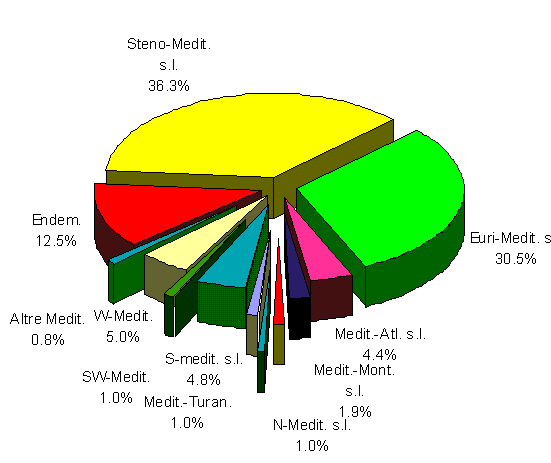
Fig. 4 and Tab. 8 report the chorological spectrum and categories grouped in macroforms for a better comparison. The second spectrum (Fig. 5 and Tab. 9) represents the main Mediterranean elements, while the third (Fig. 6 and Tab. 10) shows all the Mediterranean subelements according to biogeographic and pivotal criteria. These spectra clearly show dominance of the Mediterranean species (71.6%) and in particular of steno-Mediterranean (26%), euri-Mediterranean (21.8%) and endemic (9%) elements. The western-Mediterranean (3.5%), southern-Mediterranean (3.5%) and Atlantic-Mediterranean (3.1) components appear important in identifying the barycentre of the studied area.
Fig. 4. General chorological spectrum.
CHOROLOGICAL TYPE n. taxa
%
Cosmopolites 88
13.2
Tropical s.l. 23
3.4
Circumboreal 20
3.0
Eurasiatic s.l. 59
8.8
Mediterranean 479
71.6
Total 669
100.0
Tab. 8. General chorological types.
Fig. 5. Chorological spectrum of the main Mediterranean elements.
CHOROGICAL TYPE n. taxa
%
Endem. 60
9.0
Steno-Medit. 174
26.0
Euri-Medit. 147
21.8
Medit.-Atl. 21
3.1
Medit.-Mount. 9
1.3
S-medit. 23
3.5
W-Medit. 23
3.5
Other Medit. 22
3.3
Other non Medit. 190
28.4
Total 669
100.0
Tab. 9. Main Mediterranean chorological types.
Fig. 6. Chorological spectrum of Mediterranean subelements.
CHOROGICAL TYPE n. taxa
%
Endem. 60
12.50
Steno-Medit. s.s. 127
W-Steno-Medit. 30
Steno-Medit.-Macarones. 9
Steno-Medit.-Turan. 8
Steno-Medit. s.l. 174
36.3
Euri-Medit. s.s. 127
Euri-Medit.-Turan. 10
W-Euri-Medit. 6
Euri.-Medit.-Macarones. 4
Euri-Medit. s.l. 147
30.5
Medit.-Atl. s.s. 15
Submedit.-Subatl. 6
Medit.-Atl. s.l. 21
4.4
Medit.-Mount. s.s. 6
W-Medit.-Mount. 3
Medit.-Mount. s.l. 9
1.9
CHOROGICAL TYPE n. taxa
%
N-Medit. s.s. 3
NE-Medit. 1
NW-Medit. 1
N-Medit. s.l. 5
1.0
E-Medit. s.s. 2
E-Medit.-Turan. 1
E-Medit. s.l. 3
0.6
Medit.-Turan. 5
1.0
S-Medit. s.s. 20
S-Medit.-Turan. 3
S-Medit. s.l. 23
4.8
SW-Medit. 5
1.0
W-Medit. 19
W-Medit.-Macarones. 4
W-Medit. 23
5.0
Other Medit. 4
0.8
Total Medit. 479
100.0
Tab. 10. Mediterranean subelements.
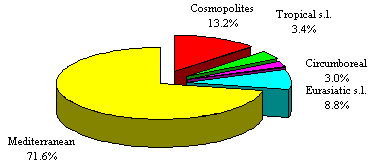
The spectrum of endemic entities (60 taxa, 51 of which specific, 7 subspecific and 2 hybrids) was made by separating the Sardinian component from the Sardo-Corsican and other widely distributed components (Fig. 7 and Tab. 11). According to Arrigoni and Di Tommaso (1991), Tyrrhenian entities and those present also on the French coasts (Ga) and on the Hy�res islands (H) have been included in the insular Tyrrhenian endemic, Tyrrhenian and western Mediterranean categories. The graph shows a clear dominance of Sardo-Corsican (31.7%) and Sardinian (21.7%) endemics, that together reach 53.4% of the total. The Sardo-Corsican and Sardo-Corso-Tuscan Archipelago figure (16.7%) is so high as a result of the siliceous substrate that tends to dominate the morphologies of the topmost zones, the rocky faces and areas above an altitude of 400-600 m.
Fig. 7. Chorological spectrum of the endemic component.
n. taxa
%
Sardinia 13
21.7
Sardinia-Corsica 19
31.7
Sardinia-Corsica-Tuscan Archipelago 10
16.7
Sardinia-Corsica-Tuscan Archipelago-Balearic Islands 6
10.0
Tyrrhenian insular endemics (sensu Arrigoni & al. op. cit.) 4
6.7
Tyrrhenian endemics (sensu Arrigoni & al. op. cit.) 3
5.0
Western-Mediterranean endemics (sensu Arrigoni & al. op. cit.) 5
8.3
Total 60
100.0
Tab. 11. Number and percentage of endemics.

The endemic component (Tab. 12) and in particular the exclusive taxa Armeria sulcitana Arrigoni, Genista valsecchiae Brullo & De Marco and Helichrysum montelinasanum E. Schmid show the floristic independence of the sector, in confirmation of what was pointed out by Mossa & al. (1996).
The observation of Borago pygmaea (DC.) Chater & Greuter, Galium corsicum Sprengel, Genista aetnensis (Biv.) DC., Genista valsecchiae Brullo & De Marco, Linaria arcusangeli Atzei & Camarda, Ophrys x maremmae O. & E. Danesch nsubsp. woodii Corrias and Orchis x penzigiana Camus nsubsp. sardoa Scrugli & Grasso contributes to give a better definition of the distribution area and of the ecology of these entities.
In particular, the finding of Borago pygmaea (DC.) Chater & Greuter in the Sa Senna Manna and Longufresu channels confirms the bibliographical datum reported by Valsecchi (op. cit.) on an exsiccata from the Capoterra area, preserved at the herbarium CAG without any information on the date or gatherer.
As regards Galium corsicum Sprengel, this is the first report in Sulcis and the second in southern Sardinia. It had only been found on Monte Linas by Angiolino and Chiappini (1983).
Genista aetnensis (Biv.) DC. had only been reported at Is Cannoneris (Arrigoni et al. 1967), while Genista valsecchiae Brullo & De Marco was believed to be coastal, since it had only been reported for south-western coasts (Brullo & De Marco 1996).
The finding of Linaria arcusangeli Atzei & Camarda both on Monte Arcosu and on Monte Lattias, extends the distribution area of this species to the Sulcis mountains , since it had only been reported at Arco dell'Angelo (Atzei & Camarda 1984, Camarda 1985) and Rio Cannas (Ballero 1988).
As regards Ophrys x maremmae O. & E. Danesch nsubsp. woodii Corrias and Orchis x penzigiana Camus nsubsp. sardoa Scrugli & Grasso, it should be mentioned that this is the first report for Sulcis and that only one specimen of each had been found.
The data confirming the presence of rare endemics such as Genista morisii Colla, Mentha requienii Bentham, Ophrys holoserica (Burm.) Greuter subsp. chestermanii Wood, Orchis mascula (L.) L. subsp. ichnusae Corrias and Soleirolia soleirolii (Req.) Dandy are also worthy of mention.
The following were also included among the endemic entities Dipsacus ferox Loisel. in agreement with the suggestion by Bocchieri & al. (op. cit.) and Helichrysum italicum (Roth) Don subsp. microphyllum (Willd.) Nyman according to a proposal by Biondi & al. (op. cit.).
However, as regards Serapias nurrica Corrias, it should be pointed out that it has not been included among endemic entities, in the light of recent findings in south Portugal (Salkowski 1993) and in that of of taxonomic and phytogeographic studies (Scrugli 1997). Based on these studies and following the proposal by Scrugli (op. cit.), it has been deemed suitable to consider this species a western steno-Mediterranean.
ENTITIES DISTRIBUTION AREA Allium parciflorum Viv. Sa-Co Arenaria balearica L. Sa-Co-AT-Bl Aristolochia rotunda L. subsp. insularis (Nardi & Arrigoni) Gamisans Sa-Co Aristolochia tyrrhena Nardi & Arrigoni Sa Armeria sulcitana Arrigoni Sa Arum pictum L. f. Sa-Co-AT-Bl Barbarea rupicola Moris Sa-Co Bellium bellidioides L. Sa-Co-Bl Bituminaria morisiana (Pign. & Metlesics) Greuter Sa-La Galite (Tn) Borago pygmaea (DC.) Chater & Greuter Sa-Co-AT Bryonia marmorata Petit Sa-Co Carex microcarpa Bertol. ex Moris Sa-Co-AT Crocus minimus DC. in R�dout� Sa-Co-AT Cymbalaria aequitriloba (Viv.) A. Chev. Sa-Co-AT-Bl Delphinium pictum Willd. Sa-Co-Bl-H Dianthus siculus C. Presl. Sa-Co-Si Dipsacus ferox Loisel. Sa-Co Euphorbia cupanii Guss. ex Bertol. Sa-Co-Si Euphorbia semiperfoliata Viv. Sa-Co Galium corsicum Sprengel Sa-Co Genista aetnensis (Biv.) DC. Sa-Si Genista corsica (Loisel.) DC. in Lam. & DC. Sa-Co Genista valsecchiae Brullo & De Marco Sa Genista morisii Colla Sa Helichrysum italicum (Roth) subsp. microphyllum (Willd.) Nyman Sa-Co-Bl Helichrysum montelinasanum E. Schmid Sa Hypericum hircinum L. Sa-Co-AT Leucanthemum flosculosum (L.) P. Giraud Sa-Co Linaria arcusangeli Atzei & Camarda Sa Mentha insularis Req. ex Gren. & Godr. Sa-Co-AT-Bl Mentha requienii Bentham Sa-Co-AT Mercurialis corsica Cosson Sa-Co Oenanthe lisae Moris Sa Ophrys holoserica (Burm.) Greuter subsp. chestermanii Wood Sa Ophrys morisii (Martelli) So� in Keller & al. Sa-Co Ophrys x maremmae O. & E. Danesch nsubsp. woodii Corrias Sa Orchis mascula (L.) L. subsp. ichnusae Corrias Sa Orchis x penzigiana Camus nsubsp. sardoa Scrugli & Grasso Sa Ornithogalum biflorum Jordan & Fourr. Sa-Co Orobanche rigens Loisel. Sa-Co Pancratium illyricum L. Sa-Co-AT Polygonum scoparium Req. ex Loisel. Sa-Co Ptilostemon casabonae (L.) Greuter Sa-Co-AT-H Ranunculus cordiger Viv. subsp. diffusus (Moris) Arrigoni Sa-Co Ranunculus revelieri Boreau Sa-Co-Ga Robertia taraxacoides (Loisel.) DC. Sa-Co-Si-It Romulea requieni Parl. Sa-Co-It Salix arrigoni Brullo Sa Saxifraga corsica Gren. & Godr. Sa-Co Scrophularia ramosissima Loisel. Sa-Co-Bl-Ga Scrophularia trifoliata L. Sa-Co-AT Seseli bocconi Guss. subsp. praecox Gamisans Sa-Co Silene nodulosa Viv. Sa-Co Soleirolia soleirolii (Req.) Dandy Sa-Co-AT-(Bl) Stachys corsica Pers. Sa-Co Stachys glutinosa L. Sa-Co-AT Teucrium marum L. Sa-Co-AT-(Bl)-H Urtica atrovirens Req. ex Loisel. Sa-Co-AT-(Bl)-It Verbascum conocarpum Moris Sa-Co-AT Viola corsica Nyman subsp. limbarae Merxm. & Lippert Sa Tab. 12. The endemic component of Rio Santa Lucia.
As regards the phytogeographically interesting taxa, the finding of Urginea fugax (Moris) Steinh. (Arrigoni 1964), Salix atrocinerea Brot. (Camarda & al. 1993) and Spiranthes aestivalis (Poiret in Lam.) Rich. (Marras & al. 1995) is confirmed.
As regards Taxus baccata L., a second population in the Senna Manna channel and a number of isolated specimens on the ridges of Monte Lattias and in Rio su Fundu are pointed out.
The following are new findings for the mountain complex of Sulcis: Blechnum spicant (L.) Roth, Cneorum tricoccon L., Laurus nobilis L., Leucojum aestivum L. subsp. pulchellum (Salisb.) Briq. and Quercus morisii Borz�.
In particular, as regards Blechnum spicant (L.) Roth, it should be pointed out that this is the third report in Sardinia and the first in Sulcis, since this entity had only been reported in Gennargentu and Limbara (Ferrarini & al. op. cit.).
Considering their inaccessible positions and large size, the populations of Laurus nobilis L. are certainly among the most interesting examples of laurisilva relics in Sardinia.
![]()
